Genes ligados ao sexo
Hoje, sabemos que os genes – “fatores” de Mendel – são segmentos de DNA localizados ao longo dos cromossomos. Podemos ver a localização de um determinado gene por meio da marcação dos cromossomos com um corante fluorescente que destaca aquele gene.
Entretanto, os “fatores de herança”, de Gregor Mendel, eram apenas um conceito abstrato quando Mendel propôs sua existência em 1860. Naquele momento, não se conhecia nenhuma estrutura celular que pudesse abrigar essas unidades imaginárias, e a maioria dos biólogos não acreditava nas leis de herança propostas por Mendel.
Com o uso de técnicas melhoradas de microscopia, os citologistas descobriram o processo da mitose em 1875 e o da meiose nos anos 1890. A citologia e a genética convergiram quando os biólogos começaram a observar os paralelos entre o comportamento dos cromossomos e o comportamento dos fatores de herança, propostos por Mendel, durante os ciclos de vida sexual.
Tanto cromossomos como genes estão presentes aos pares nas células diploides; os cromossomos homólogos se separam e os alelos segregam durante o processo de meiose; a fertilização restabelece a condição pareada tanto para os cromossomos quanto para os genes.
Por volta de 1902, Walter S. Sutton, Theodor Boveri e outros observaram de forma independente esses paralelos, e a teoria cromossômica da herança começou a ser elaborada. De acordo com essa teoria, os genes mendelianos têm loci (posições) específicos nos cromossomos e são os cromossomos que sofrem a segregação e a segregação independente.
Morgan mostrou que a herança mendeliana tem sua base física no comportamento dos cromossomos
A primeira evidência sólida que associa um determinado gene a um determinado cromossomo apareceu no início do século XX, a partir do trabalho de Thomas Hunt Morgan, embriologista experimental da Universidade da Columbia.
Embora Morgan inicialmente não acreditasse tanto sobre a genética de Mendel como sobre a teoria dos cromossomos, seus experimentos iniciais forneceram evidências convincentes de que os cromossomos realmente são o local dos fatores de herança de Mendel.
Organismo experimental escolhido por Morgan
Para seu trabalho, Morgan selecionou uma espécie de mosca-das-frutas, Drosophila melanogaster, inseto comum que se alimenta de fungos que crescem nas frutas. As moscas-das-frutas são reprodutoras prolíficas; um único cruzamento produz centenas de descendentes e uma nova geração pode ser produzida a cada duas semanas.
Outra vantagem da mosca-das-frutas é que ela possui apenas quatro pares de cromossomos, facilmente distinguíveis com um microscópio óptico. Existem três pares de autossomos e um par de cromossomos sexuais. Moscas-das-frutas fêmeas têm um par de cromossomos X homólogos, e os machos, um cromossomo X e um cromossomo Y.
Morgan e seus estudantes inventaram uma notação para simbolizar os alelos de Drosophila, ainda amplamente utilizada para mosca-das-frutas. Para um dado caractere em moscas, o gene toma o símbolo a partir do primeiro mutante descoberto (tipo não selvagem).
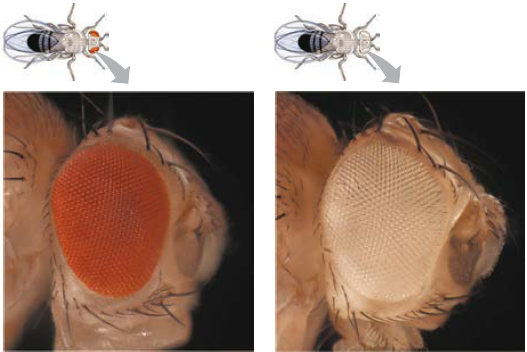
Assim, o alelo para olhos brancos em Drosophila é simbolizado por w. Um + sobrescrito identifica o alelo para a característica tipo selvagem; w+ para o alelo dos olhos vermelhos, por exemplo.
Experimento
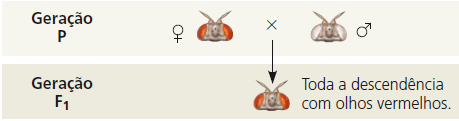
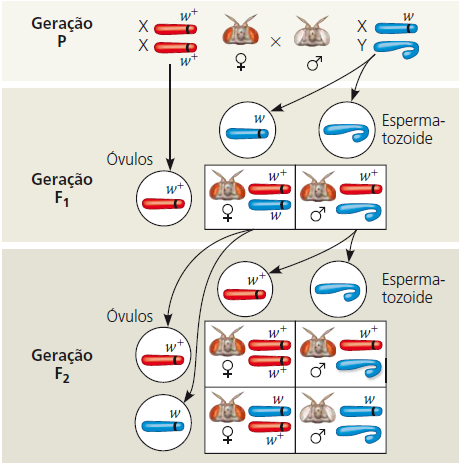
Thomas Hunt Morgan queria analisar o comportamento de dois alelos de um gene para cor dos olhos da mosca-das-frutas. Em cruzamentos semelhantes àqueles realizados por Mendel com ervilhas, Morgan e colaboradores cruzaram uma fêmea do tipo selvagem (olhos vermelhos) com um macho mutante de olhos brancos.
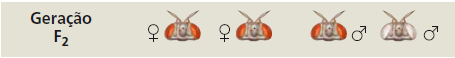
Então Morgan cruzou uma fêmea F1 de olhos vermelhos com um macho F1 de olhos vermelhos para produzir a geração F2. Por sua vez a geração F2 mostrou uma proporção mendeliana típica de 3 moscas de olhos vermelhos e 1 mosca de olhos brancos. Todavia, todas as moscas de olhos brancos eram machos; nenhuma fêmea apresentou a característica de olhos brancos.
Toda a descendência F1 apresentou olhos vermelhos; por isso, a característica mutante de olhos brancos (w) deve ser recessiva em relação à característica de olhos vermelhos tipo selvagem (w+). Uma vez que a característica recessiva – olhos brancos – foi expressada apenas em machos na geração F2, Morgan deduziu que esse gene para cor dos olhos está localizado no cromossomo X, e que não existe locus correspondente no cromossomo Y.
A base cromossômica do sexo
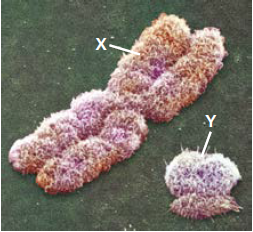
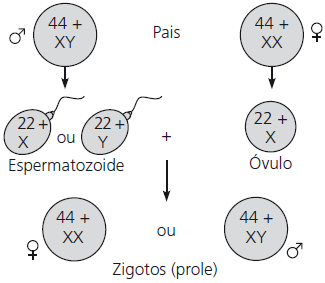
Embora as diferenças anatômicas e fisiológicas entre mulheres e homens sejam numerosas, a base cromossômica para determinação do sexo é simples. Em seres humanos e outros mamíferos, existem duas variedades de cromossomos sexuais, denominados X e Y. O cromossomo Y é muito menor do que o cromossomo X. Uma pessoa que herda dois cromossomos X, um a partir de cada progenitor, apresenta o sexo feminino; já o sexo masculino herda um cromossomo X e um cromossomo Y.
Tanto nos testículos como nos ovários, os dois cromossomos sexuais segregam durante a meiose. Cada óvulo recebe um cromossomo X. Em contrapartida, os espermatozoides se dividem em duas categorias: metade das células espermáticas que um homem produz recebe um cromossomo X e a outra metade recebe um cromossomo Y.
Herança de genes ligados ao X
Em seres humanos, os genes ligados ao X seguem o mesmo padrão de herança que Morgan observou para o locus da cor dos olhos estudado em Drosophila. Os pais passam os alelos ligados ao X para todas as filhas, mas para nenhum dos filhos. Ao contrário, as mães podem passar os alelos ligados ao X tanto para filhos como para filhas
Se uma característica ligada ao X decorre de um alelo recessivo, uma mulher apenas expressará o fenótipo se for homozigota para aquele alelo. Como os homens têm apenas um locus, qualquer um que receba o alelo recessivo a partir de sua mãe expressará a característica. Por essa razão, muito mais homens do que mulheres têm distúrbios recessivos ligados ao X.
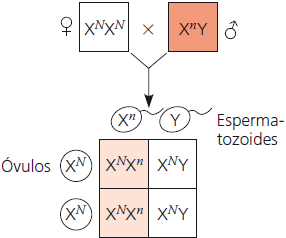
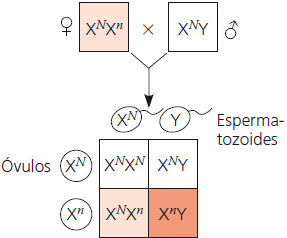
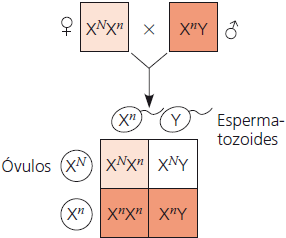
Daltonismo
Nas imagens abaixo, o daltonismo é utilizado como exemplo. O N sobrescrito representa o alelo dominante para visão normal carregado pelo cromossomo X, e o n sobrescrito representa o alelo recessivo, que possui uma mutação que causa o daltonismo. Os quadrados brancos indicam os indivíduos não afetados, os quadrados em cor de laranja claro indicam os carreadores, e os em cor de laranja escuro, os indivíduos daltônicos.
Inativação do X em fêmeas de mamíferos
Uma vez que fêmeas de mamíferos, incluindo seres humanos, herdam dois cromossomos X – o dobro do número herdado pelos machos -, você poderia se perguntar se as fêmeas produzem o dobro das proteínas codificadas pelos genes ligados ao X, comparado às quantidades nos machos.
Na verdade, um cromossomo X em cada célula nas fêmeas torna-se quase completamente inativado durante o desenvolvimento embrionário. Como resultado, as células das fêmeas e dos machos têm a mesma dose efetiva (uma cópia) da maioria dos genes ligados ao X.
O X inativo em cada célula de fêmea condensa na forma de um objeto compacto chamado de corpúsculo de Barr (descoberto pelo anatomista canadense Murray Barr), que se encontra dentro do envelope nuclear.
A geneticista britânica Mary Lyon demonstrou que a seleção de qual cromossomo X formará um corpúsculo de Barr ocorre de forma aleatória e independente em cada célula embrionária presente no momento da inativação do X. Como consequência, as fêmeas consistem em um mosaico de dois tipos de células: aquelas com o X ativo derivado do pai e aquelas com o X ativo derivado da mãe.
Depois que um cromossomo X é inativado em determinada célula, todos os descendentes mitóticos daquela célula possuem o mesmo X inativo. Assim, se uma fêmea é heterozigota para uma característica ligada ao sexo, cerca da metade das suas células expressarão um alelo, ao passo que as outras expressarão o alelo alternativo.
A figura abaixo mostra como essa formação de mosaicos resulta na coloração matizada de um gato com pelagem tartaruga.

Em seres humanos, a formação de mosaicos pode ser observada em uma mutação recessiva ligada ao X que previne o desenvolvimento das glândulas sudoríparas. Uma mulher heterozigota para essa característica tem áreas de pele normal e áreas de pele sem glândulas sudoríparas.
