Estruturas celulares
No decorrer dos séculos XIX e XX, a melhora da microscopia revelou mais complexidade nas células. Foram encontradas as primeiras organelas (estruturas dentro da célula), embora nem sempre sua função fosse compreendida na hora. Na verdade, as organelas são fundamentais. Algumas executam os processos bioquímicos da vida, como metabolizar os alimentos (seja qual for a fonte), liberar e armazenar energia.
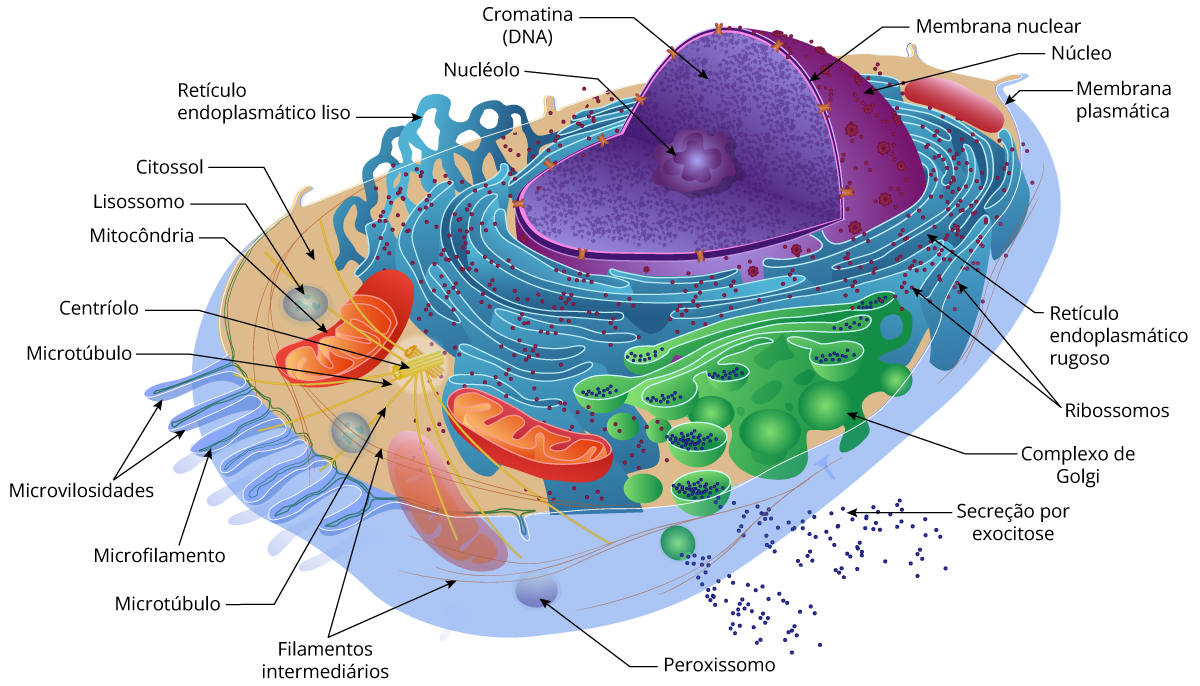
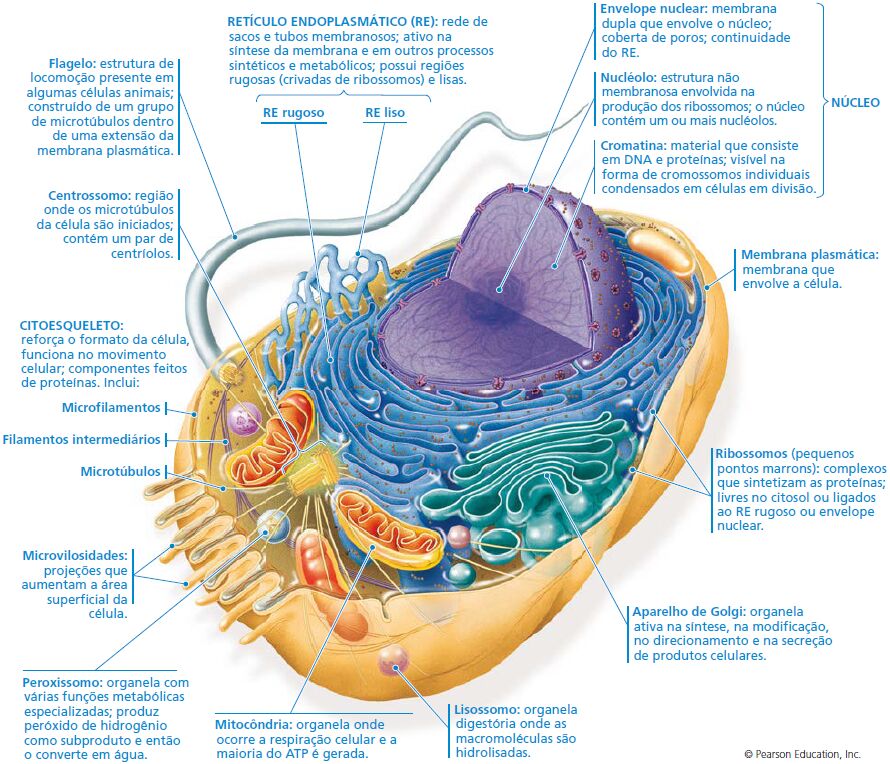
Panorama das células eucarióticas
Além da membrana plasmática na superfície externa, a célula eucariótica possui membranas internas arranjadas de forma elaborada, que dividem a célula em compartimentos – as organelas antes mencionadas. Os compartimentos celulares fornecem meios diferentes que facilitam as funções metabólicas específicas, de modo que processos incompatíveis possam ocorrer simultaneamente dentro de uma única célula.
Célula animal
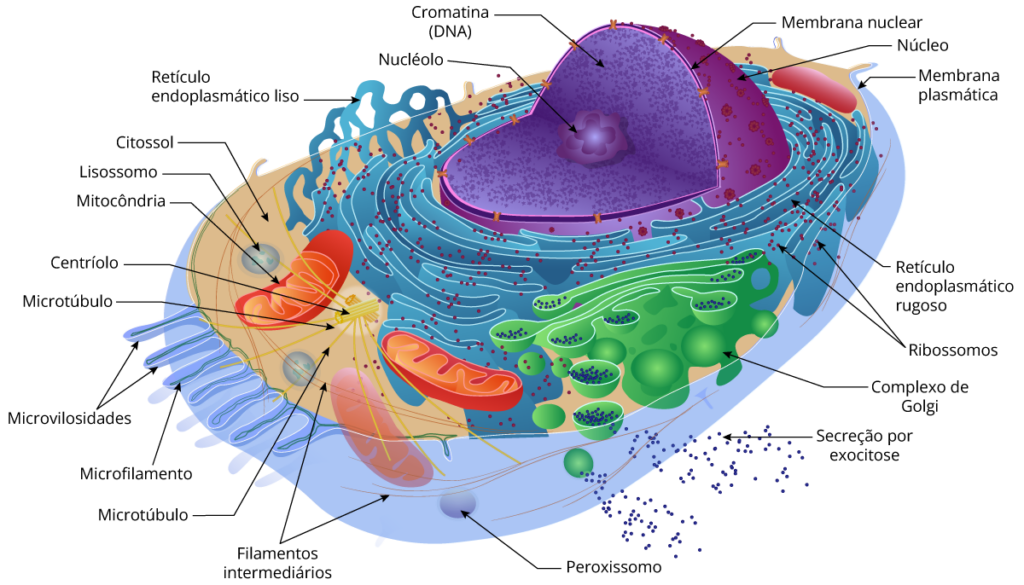
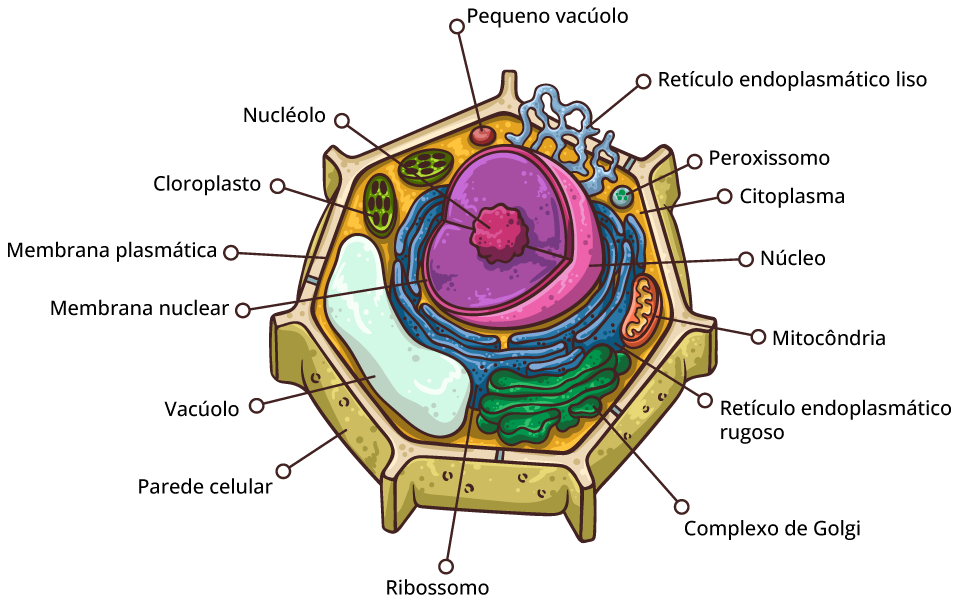
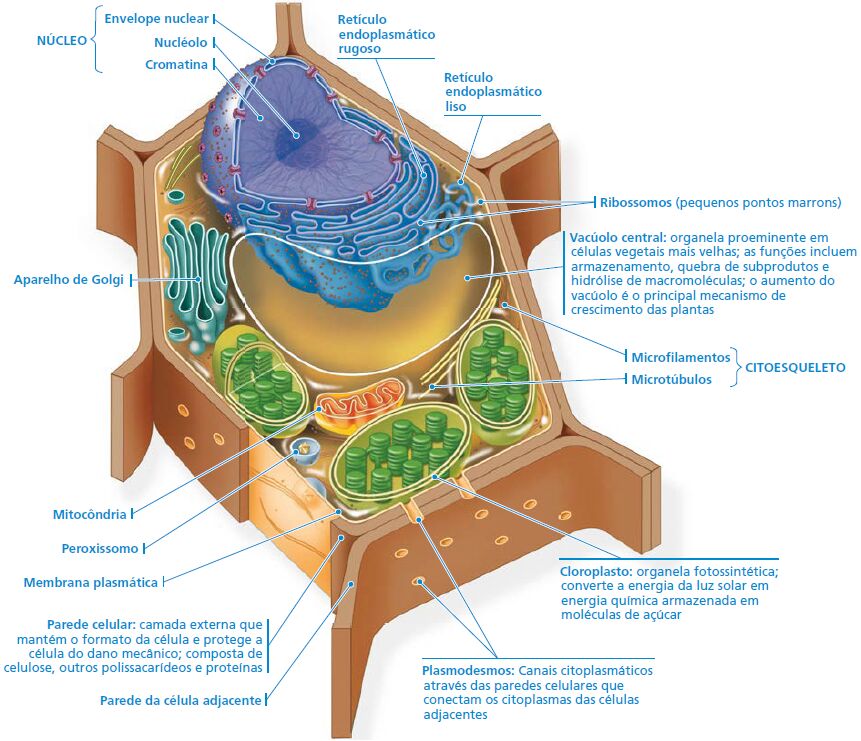
Célula animal x célula vegetal
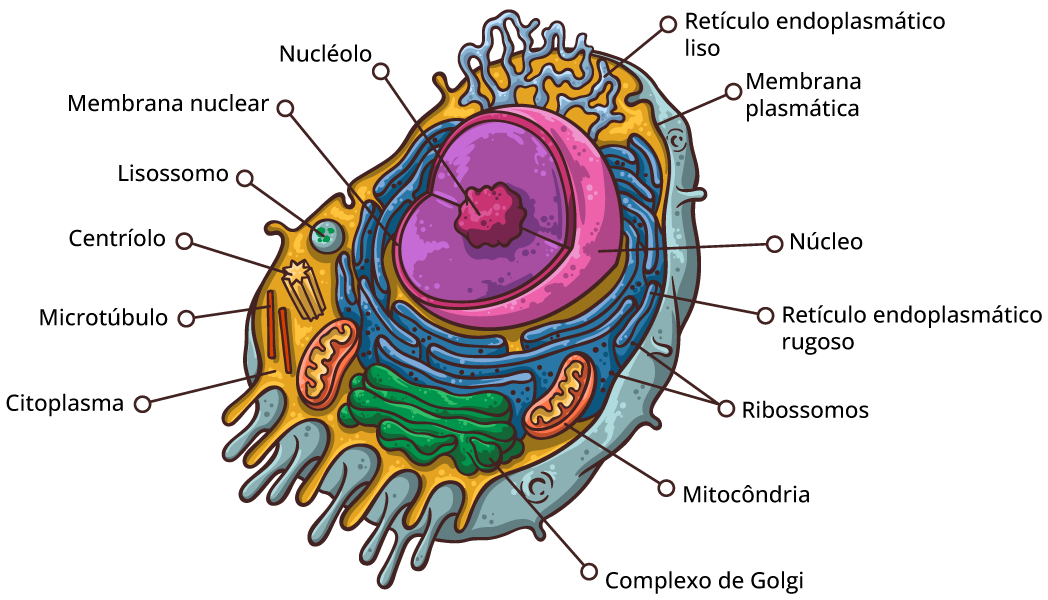
Comparação entre células procarióticas e eucarióticas
Todas as células compartilham certas características básicas: são ligadas por uma barreira seletiva chamada de membrana plasmática. Dentro de todas as células existe um semifluido, substância semelhante à gelatina chamada de citosol, na qual os componentes subcelulares estão suspensos. Todas as células contêm cromossomos, que carregam os genes na forma de DNA. E todas as células contêm ribossomos, minúsculos complexos que sintetizam as proteínas de acordo com as instruções a partir dos genes.
A principal diferença entre células procarióticas e eucarióticas é a localização do seu DNA. Na célula eucariótica, a maioria do DNA está na organela chamada de núcleo, ligada por uma membrana dupla. Na célula procariótica o DNA está concentrado em uma região não envolta por membrana, chamada de nucleoide. Eucariótico significa “com núcleo verdadeiro”(do grego, eu, verdadeiro, e karyon, cerne, referindo-se ao núcleo), enquanto procariótico significa “antes do núcleo” (do grego, pro, antes), refletindo a evolução das células procarióticas.
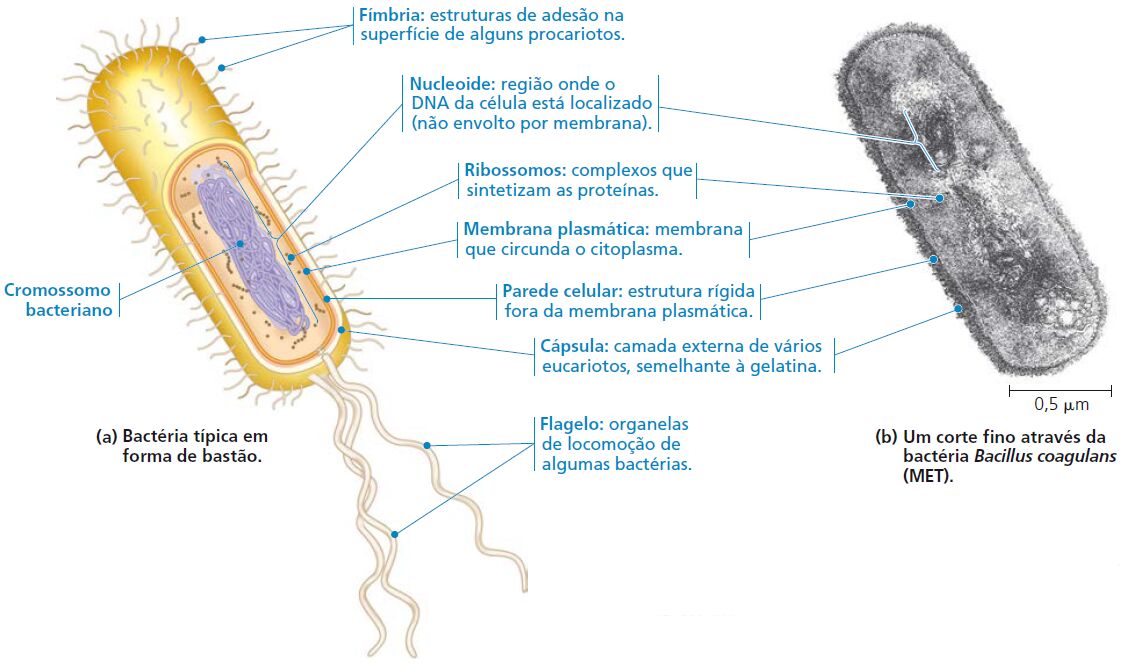
Célula procariota x célula eucariota
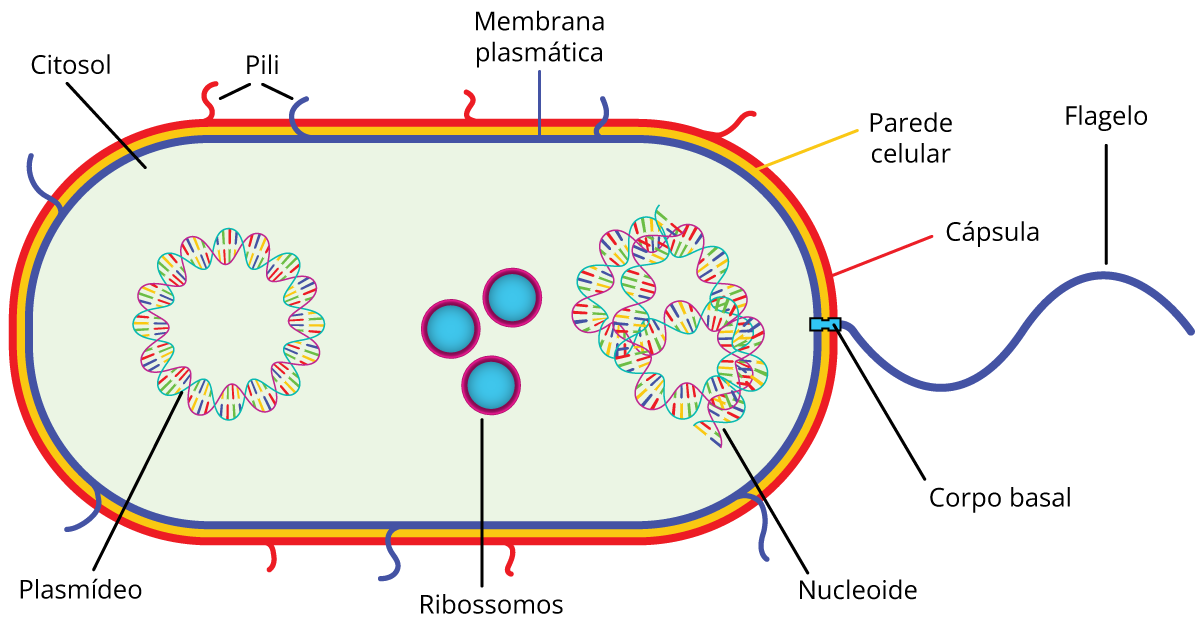
- Pili ou fímbrias: presente em algumas bactérias, serve para fixá-las ao substrato.
- Cápsula: estrutura de proteção de algumas bactérias.
- Plasmídeo: fragmento de DNA.
A poderosa mitocôndria
A organela mais importante é a mitocôndria, geralmente chamada de “usina de energia” da célula por ser responsável pela produção da maior parte do ATP (trifosfato de adenosina) que as células usam para armazenar energia química. Provavelmente, a mitocôndria foi observada pela primeira vez em na década de 1840, mas só se estabeleceu como organela em 1894 com o patologista alemão Richard Altmann, que a chamou de “bioblasto”.
O anatomista americano Benjamin F. Kingsbury foi o primeiro a ligar a mitocôndria à respiração celular em 1912, mas só em 1925 a sequência envolvida foi descoberta. A complexidade do processo de respiração celular continuou a se revelar durante o século XX.
ATP: a moeda energética dos seres vivos
Estruturalmente, o ATP consiste em três grupos fosfatos ligados à adenosina, composta de adenina e ribose. A reação de transferência de energia ocorre por meio de hidrólise: a remoção do último grupo fosfato (P) por adição de uma molécula de água, liberando assim, energia. Como resultado tem-se uma molécula de ADP (difosfato de adenosina).
A energia liberada pela decomposição do ATP é constantemente utilizada pela célula. Contudo, o ATP pode ser reconstituído por meio da enzima ATP sintase, que promove a adição de um grupo fosfato (P) no ADP.
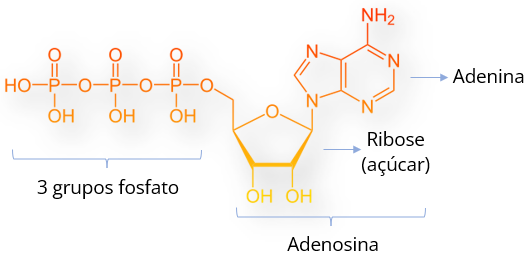
O ATP contém a energia necessária para manter as atividades das células. Esta energia é liberada quando a ligação do último grupo fosfato é quebrada. As nossas células conseguem sintetizar o ATP, refazendo a ligação de um fosfato no ADP. Este processo acontece no interior das usinas das células, as mitocôndrias.

Os cloroplastos
Organela responsável por um dos processos mais importantes do planeta: a fotossíntese. Esse processo é a via pela qual praticamente toda a energia entra em nossa biosfera. A cada ano, mais de 250 milhões de toneladas de açúcar são produzidas na natureza pelos organismos fotossintetizantes. Sem esse fluxo de energia a partir do Sol e canalizado em grande parte pelos cloroplastos das células eucarióticas, o ritmo da vida neste planeta seria rapidamente diminuído e, então, praticamente cessaria por inteiro.
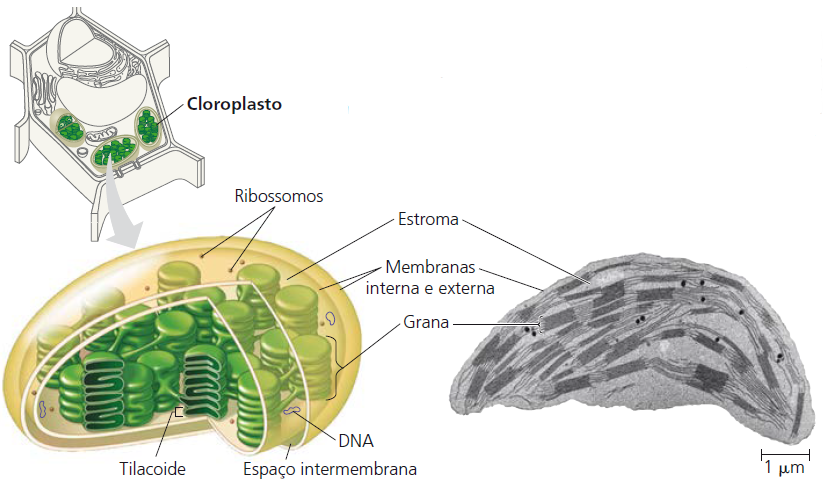
As origens evolutivas de mitocôndrias e cloroplastos
As mitocôndrias e os cloroplastos apresentam semelhanças com bactérias que levam à teoria endossimbionte. Essa teoria estabelece que um ancestral das células eucarióticas engolfou uma célula procariótica que utilizava oxigênio e não realizava fotossíntese. Por fim, a célula engolfada formou um relacionamento com a célula hospedeira se tornando uma endossimbionte (uma célula vivendo dentro da outra célula.)
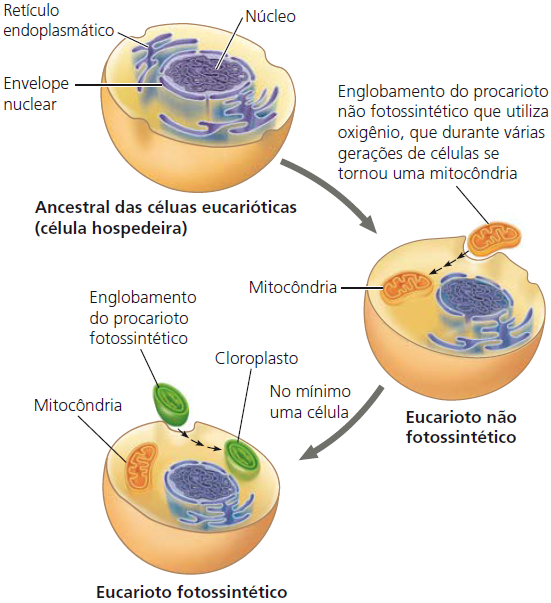
Ainda, durante o curso da evolução, a célula hospedeira e sua endossimbionte fundiram-se em um único organismo, uma célula eucariótica com uma mitocôndria. Pelo menos uma dessas células pode ter captado um procarioto fotossintético, tornando-se o ancestral das células eucarióticas que contêm cloroplastos. Essa teoria é consistente com muitas características estruturais de mitocôndrias e cloroplastos.
Primeiro, em vez de estarem envoltos por uma membrana simples como as organelas do sistema de endomembranas, mitocôndrias e os cloroplastos típicos têm duas membranas ao seu redor. Segundo, assim como os procariotos, as mitocôndrias e cloroplastos contêm ribossomos, além de moléculas de DNA circular associadas com suas membranas internas. Terceiro, também consistente com suas prováveis origens evolutivas, as mitocôndrias e os cloroplastos são organelas autônomas (um pouco independentes) que crescem e se reproduzem dentro da célula.
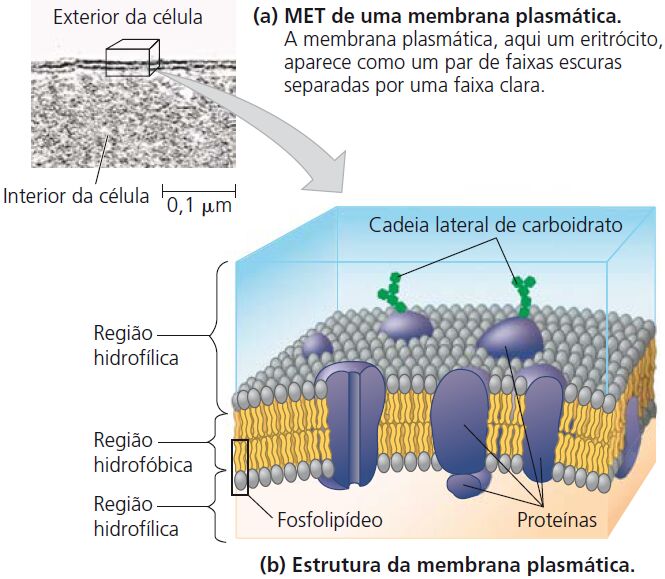
A membrana plasmática
Forma a superfície externa flexível da célula, separando o ambiente interno da célula de seu ambiente externo. Regula o fluxo de materiais para dentro e para fora da célula, para manter um ambiente apropriado às atividades celulares normais. A membrana também exerce uma função essencial na comunicação entre células e entre as células e seu ambiente externo. A principal molécula que forma a membrana é o fosfolipídio.
O citoplasma
Consiste em todos os conteúdos celulares entre a membrana plasmática e o núcleo. O citoplasma é dividido em dois componentes: citosol e organelas. Citosol é a porção líquida do citoplasma, que consiste principalmente em água mais solutos dissolvidos e partículas suspensas. Também é chamado de fluido intracelular. No citosol existem vários tipos diferentes de organelas, cada uma das quais possui uma estrutura característica e funções específicas.
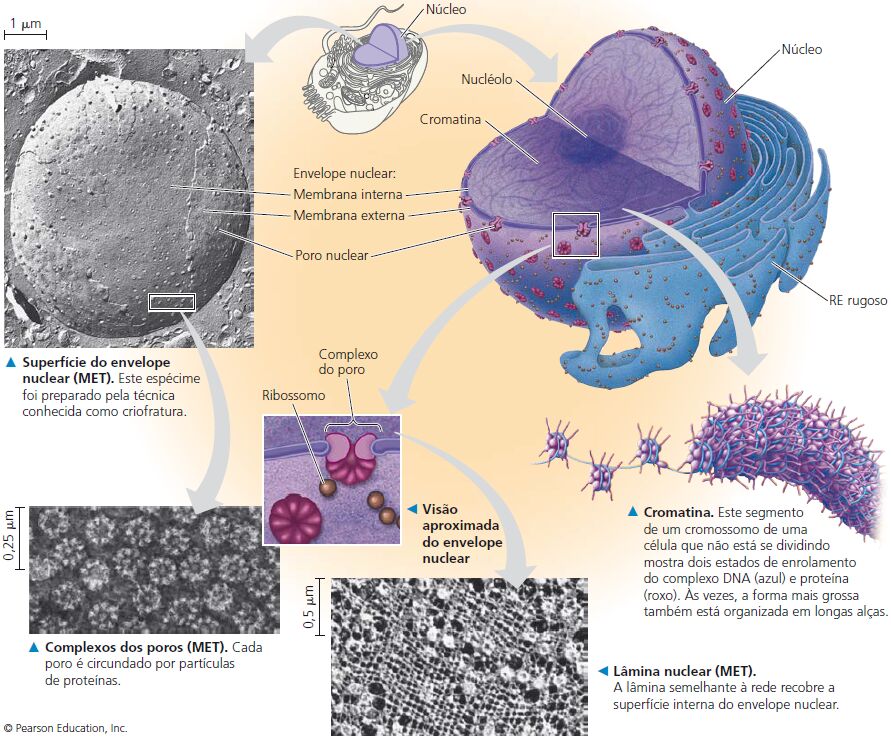
O núcleo
O núcleo contém a maioria dos genes na célula eucariótica (alguns genes estão localizados nas mitocôndrias e nos cloroplastos). Ele geralmente é a organela mais evidente, medindo cerca de 5 μm de diâmetro. O envelope nuclear envolve o núcleo, separando seu conteúdo do citoplasma.
O envelope nuclear é uma membrana dupla. Cada uma das duas membranas é uma bicamada lipídica com proteínas associadas, separadas por um espaço de 20 a 40 nm. O envelope é perfurado por estruturas de poros que medem cerca de 100 nm de diâmetro. O lado nuclear do envelope é revestido pela lâmina nuclear, um arranjo de filamentos proteicos, semelhante à rede, que mantém a forma do núcleo dando suporte mecânico do envelope nuclear.
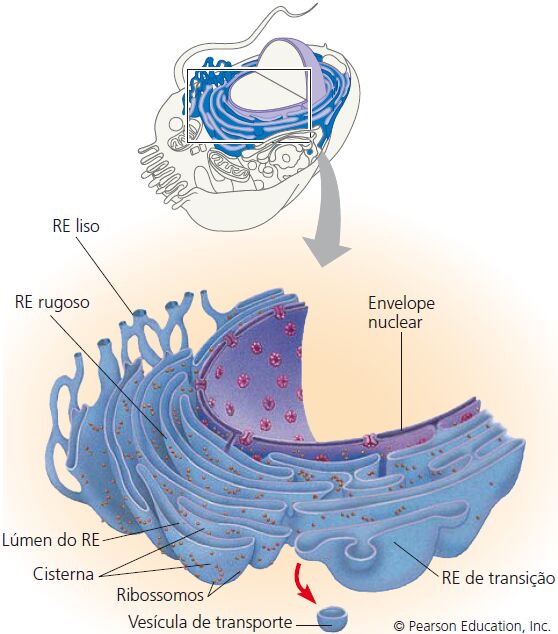
O retículo endoplasmático
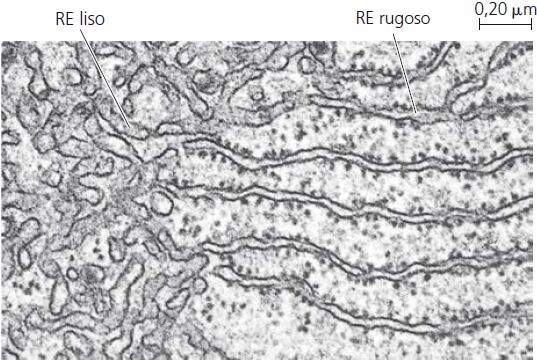
É uma rede de membranas pregueadas na forma de sacos ou túbulos achatados. O retículo se estende por todo o citoplasma e é tão extenso que constitui mais da metade das superfícies membranáceas no interior do citoplasma da maioria das células. As células contém duas formas distintas de retículo endoplasmático (RE), que diferem em estrutura e função. O RE rugoso se estende a partir do envoltório nuclear (membrana em torno do núcleo) e tem aparência “rugosa”, porque sua superfície externa é cravejada de ribossomos. As proteínas sintetizadas pelos ribossomos fixados no RE rugoso entram nos espaços internos do RE, para processamento e classificação.
O RE liso se estende a partir do RE rugoso para formar uma rede de túbulos membranáceos. Como você já deve ter concluído, o RE liso possui uma aparência “lisa” pela falta de ribossomos. O RE liso é o local de síntese dos ácidos graxos e esteroides, como os estrogênios e a testosterona. Nas células do fígado, enzimas do RE liso também ajudam a liberar glicose na corrente sanguínea, inativar e desintoxicar uma variedade de fármacos e substâncias potencialmente prejudiciais, incluindo álcool, pesticidas e carcinógenos (agentes produtores de câncer). Nas células musculares, íons cálcio necessários à contração muscular são armazenados e liberados a partir de uma forma de RE liso chamada retículo sarcoplasmático.
O complexo de Golgi
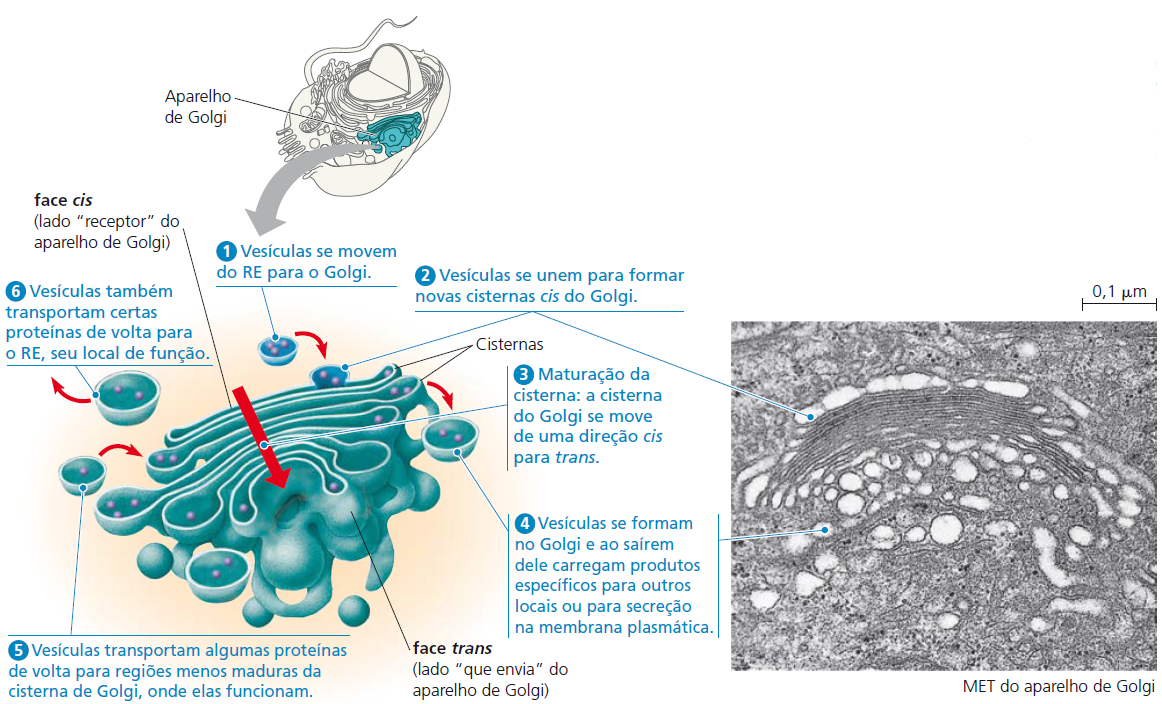
Após as proteínas serem sintetizadas em um ribossomo ligado ao RE rugoso, a maioria geralmente é transportada para outra região da célula. O primeiro passo nessa via de transporte é por meio de uma organela denominada complexo de Golgi. Este consiste de 3 a 20 cisternas, sacos membranáceos achatados com margens protuberantes, que se assemelham a uma pilha de pães sírios.
As proteínas sintetizadas pelo RE rugoso, entram no complexo de Golgi e são modificadas para formar glicoproteínas e lipoproteínas. Em seguida são classificadas e acondicionadas em vesículas. A maioria das células possuem vários complexos de Golgi. O complexo de Golgi é mais extenso nas células que secretam proteínas. Portanto, ele modifica, classifica, acondiciona e transporta proteínas recebidas pelo RE rugoso, além de formar vesículas secretoras que liberam proteínas processadas via exocitose no líquido extracelular.
Os lisossomos
São vesículas revestidas por membranas, que podem conter até 60 enzimas digestivas diferentes; estas enzimas decompõem uma ampla variedade de moléculas, uma vez que o lisossomo se funde com vesículas formadas durante a endocitose. A membrana lisossômica contém proteínas transportadoras que permitem aos produtos finais da digestão, como monossacarídeos, ácidos graxos e aminoácidos, serem transportados para o citosol.
As enzimas lisossômicas também ajudam a reciclar as estruturas desgastadas. Um lisossomo engolfa outra organela, digerindo-a e retornando os componentes digeridos ao citosol para reutilização. Dessa forma, organelas senis são continuamente substituídas. o processo pelo qual organelas desgastadas são digeridas é chamado autofagia.
Um hepatócito humano (célula do fígado), por exemplo, recicla aproximadamente metade do seu conteúdo a cada semana. Enzimas lisossômicas podem destruir completamente a célula, um processo conhecido como autólise. A autólise ocorre em algumas condições patológicas e também é responsável pela deterioração tecidual que ocorre logo após a morte.
Os peroxissomos
Outro grupo de organelas semelhantes em estrutura aos lisossomos, mas menores, são chamados peroxissomos. Os peroxissomos contêm diversas oxidases, enzimas que oxidam (removem átomos de hidrogênio) várias substâncias orgânicas. Por exemplo, os aminoácidos e os ácidos graxos são oxidados nos peroxissomos, como parte do metabolismo normal.
Além disso, as enzimas nos peroxissomos também oxidam substâncias tóxicas. Portanto, são muito abundantes no fígado, local em que ocorre a desintoxicação de álcool e de outras substâncias prejudiciais. Um subproduto das reações de oxidação é o peróxido de hidrogênio (H2O2), um composto potencialmente tóxico e associado a radicais livres, como o superóxido. Contudo, os peroxissomos também contêm uma enzima, denominada catalase, que decompõe o H2O2.
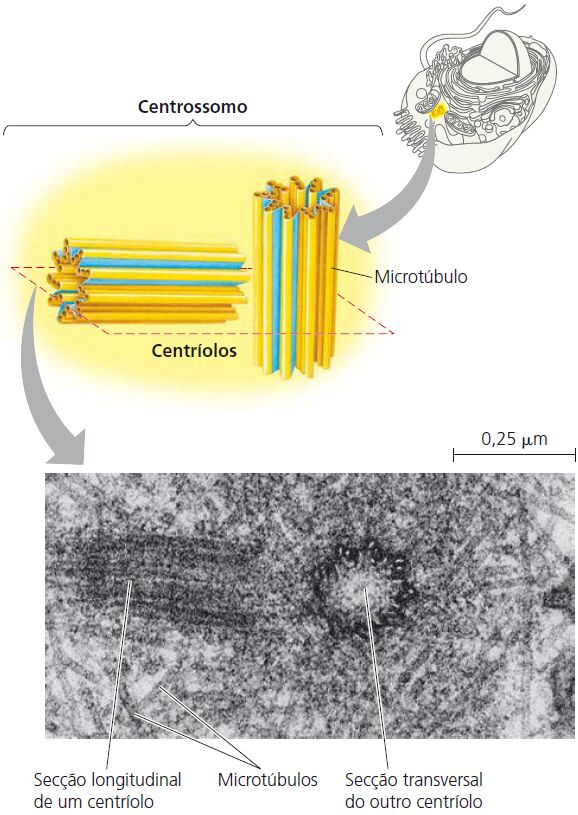
O centrossomo
Localizado próximo ao núcleo, onde encontramos os centríolos. Os centríolos são estruturas cilíndricas, cada uma composta por nove conjunto de microtúbulos dispostos em um padrão circular. Em torno dos centríolos está o material pericentriolar, que contém centenas de proteínas anulares chamadas tubulinas. As tubulinas são os centros organizadores para o desenvolvimento do fuso mitótico, que exerce uma função essencial na divisão celular. Ainda, os microtúbulos são os principais componentes estruturais e funcionais dos cílios e flagelos, ambos os quais são projeções móveis da superfície celular.
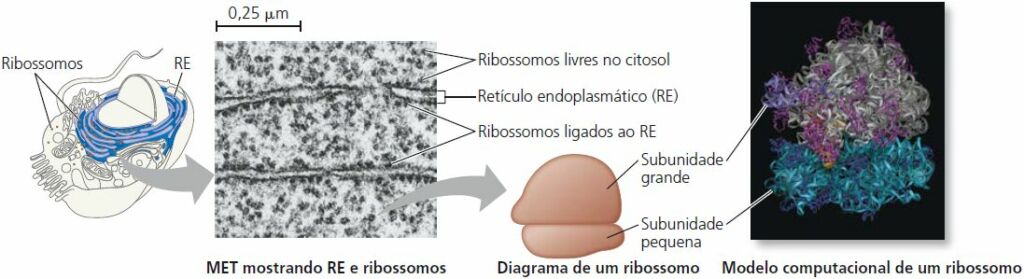
Ribossomos: fábricas de proteínas
Os ribossomos, complexos formados por RNA ribossomal e proteína, são os componentes celulares que realizam a síntese de proteínas. (Observe que os ribossomos não estão ligados a membranas e, portanto, não são considerados organelas.) Células com altas taxas de síntese proteica têm um número particularmente grande de ribossomos.
Por exemplo, uma célula do pâncreas humano, que produz muitas enzimas digestivas, tem alguns milhões de ribossomos. Não surpreendentemente, as células ativas na síntese proteica também têm nucléolos proeminentes.
Os ribossomos constroem proteínas em dois locais do citoplasma. A qualquer momento, ribossomos livres estão suspensos no citosol enquanto ribossomos ligados estão presos ao lado externo do retículo endoplasmático e do envelope nuclear. Os ribossomos ligados e os livres são estruturalmente idênticos e podem alternar os dois papéis.
Funções do citoesqueleto: suporte e motilidade
A função mais óbvia do citoesqueleto é dar sustentação mecânica à célula e manter sua forma. Isso é especialmente importante para células animais, que não possuem paredes. As incríveis força e elasticidade do citoesqueleto como um todo são baseadas na sua arquitetura. Como uma abóbada, o citoesqueleto é estabilizado pelo equilíbrio entre as forças contrárias exercidas por seus elementos.
Assim como o esqueleto de um animal ajuda a fixar as posições de outras partes do corpo, o citoesqueleto fornece âncora para várias organelas e até mesmo moléculas de enzimas citosólicas. Entretanto, o citoesqueleto é mais dinâmico do que o esqueleto animal. Ele pode ser decomposto rapidamente em uma parte da célula e remontado em uma posição nova, mudando o formato da célula.
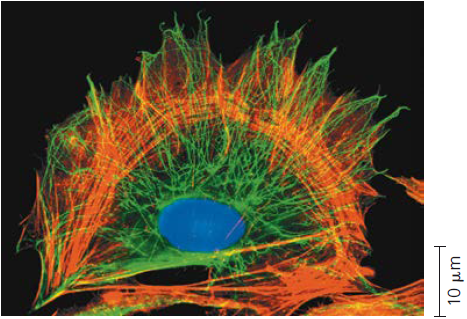
O citoesqueleto. Como mostrado nesta micrografia de fluorescência, o citoesqueleto se estende pela célula. Os elementos citoesqueléticos foram marcados com diferentes moléculas fluorescentes: verde para microtúbulos e vermelho alaranjado para os microfilamentos. Um terceiro componente do citoesqueleto, os filamentos intermediários, não estão evidentes aqui. (A cor azul marca o DNA no núcleo.)

{kind=link}
{kind=link}
{kind=link}